Механизм мышечного сокращения и расслабления биохимия. Биохимия мышц, мышечного сокращения и расслабления
по предмету
«Биохимия»
«Биохимия мышечного сокращения»
Выполнил: студент 3 курса ЕГФ
отделения «Валеология», гр. 1А
Литвиченко Е.М.
Проверил: Сайкович Е.Г.
г. Новосибирск 2000 г.
Интерес биохимии к процессам происходящим в сокращающихся мышцах основан не только на выяснении механизмов мышечных болезней, но и что может быть даже более важным – это раскрытие механизма превращения электрической энергии в механическую, минуя сложные механизмы тяг и передач.
Для того, чтобы понять механизм и биохимические процессы происходящие в сокращающихся мышцах, необходимо заглянуть в строение мышечного волокна. Структурной единицей мышечного волокна являются Миофибриллы – особым образом организованные пучки белков, располагающиеся вдоль клетки. Миофибриллы в свою очередь построены из белковых нитей (филаментов) двух типов – толстых и тонких. Основным белком толстых нитей является миозин , а тонких – актин . Миозиновые и актиновые нити – главный компонент всех сократительных систем в организме. Электронно-микроскопическое изучение показало строго упорядоченное расположение миозиновых и актиновых нитей в миофибрилле. Функциональной единицей миофибриллы является саркомер – участок миофибриллы между двумя Z-пластинками. Саркомер включает в себя пучок миозиновых нитей, серединой сцепленных по так называемой М-пластине, и проходящих между ними волокон актиновых нитей, которые в свою очередь прикреплены к Z-пластинам.
Сокращение происходит путем скольжения тонких актиновых и толстых миозиновых нитей навстречу друг другу или вдвигания актиновых нитей между миозиновыми в направлении М-линии. Максимальное укорочение достигается тогда, когда Z-пластинки, к которым прикреплены актиновые нити, приближаются к концам миозиновых нитей. При сокращении саркомер укорачивается на 25-50 %.
Саркоплазма, вмещающая миофибриллы, пронизана между ними сетью цистерн и трубочек эндоплазматического ретикулума, а также системой поперечных трубочек, которые тесно контактируют с ним, но не сообщаются.
Строение миозиновых нитей.
Миозиновые нити образованы белком миозином, молекула которого содержит две идентичные тяжелые полипептидные цепи с молекулярной массой около 200 000 и четыре легкие цепи (около 20 000). Каждая тяжелая цепь на большей части своей длины имеет конформацию a-спирали, и обе тяжелые цепи скручены между собой, образуя часть молекулы в форме палочки. С противоположных концов каждой цепи присоединены по две легкие цепи, вместе с глобулярной формой этих концов цепи они образуют «головки» молекул. Палочкообразные концы молекул могут соединяться друг с другом продольно, образуя пучки, головки молекул при этом располагаются кнаружи от пучка по спирали. Кроме того, в области М-линии пучки соединяются между собой «хвост в хвост». Каждая миозиновая нить содержит около 400 молекул миозина.
молекулы актина
молекулы тропонина молекулы тропомиозина
Другой белок, входящий в актиновые нити – тропомиозин – имеет форму палочек, он располагается вблизи желобков спиральной ленты фибриллярного актина, вдоль нее. Размер его в длину в 8 раз больше размера глобулярного актина, потому одна молекула тропомиозина контактирует сразу с семью молекулами актина и концами связаны друг с другом, образуя третью продольную спирально закрученную цепочку.
Третий белок актиновых нитей – тропонин – состоит из трех разных субъединиц и имеет глобулярную форму. Он нековалентно связан и с актином и тропомиозином таким образом, что на одну молекулу тропонина приходится одна молекула тропомиозина, кроме того одна из его субъединиц содержит Ca- связывающие центры. Тонкие актиновые нити прикреплены к Z-пластинам, тоже белковым структурам.
Механизм сокращения мышцы.
Сокращение мышц есть результат укорочения каждого саркомера, максимальное укорочение саркомера достигается тогда, когда Z-пластинки, к которым прикреплены актиновые нити, приближаются вплотную к концам миозиновых нитей.
В сокращении мышц у актиновых и миозиновых нитей свои роли: миозиновые нити содержат активный центр для гидролиза АТФ, устройство для превращения энергии АТФ в механическую энергию, устройство для сцепления с актиновыми нитями и устройства для восприятия регуляторных сигналов со стороны актиновых нитей, актиновые нити имеют механизм сцепления с миозиновыми нитями и механизм регуляции сокращения и расслабления.
Сокращение мышцы включается потенциалом действия нервного волокна, который через нервно-мышечный синапс при посредстве медиатора трансформируется в потенциал действия сарколеммы и трубочек Т-системы. Ответвления трубочек окружают каждую миофибриллу и контактируют с цистернами саркоплазматического ретикулума. В цистернах в значительной концентрации содержится Ca . Потенциал действия, поступающий по трубочкам, вызывает высвобождение ионов Ca 2+ из цистерн саркоплазматического ретикулума. Ионы Ca 2+ присоединяются к Сa-связывающей субъединице тропонина. В присутствии ионов Ca 2+ на мономерах актиновых нитей открываются центры связывания миозиновых головок, причем по всей системе тропонин – тропомиозин – актин. Как результат этих изменений – миозиновая головка присоединяется к ближайшему мономеру актина.
Головки миозина обладают высоким сродством к АТФ, так что в мышце большинство головок содержит связанный АТФ. Присоединение головки миозина к актину, активирует АТФ-азный центр, АТФ гидролизуется, АДФ и фосфат покидают активный центр, что приводит к изменению конформации миозина: возникает дополнительное напряжение, стремящееся уменьшить угол между головкой и хвостом молекулы миозина, т.е. наклонить головку в направлении М-линии. Поскольку миозиновая головка соединена с актиновой нитью, то, наклоняясь в сторону М-линии она смещает в этом же направлении и актиновую нить.
АДФ, высвобождаемые с множества головок проходят следующую трансформацию:
2 АДФ ® АТФ + АМФ
Освобожденные от АТФ головки снова притягивают к себе АТФ в связи с его высоким сродство, о чем уже упоминалось выше, присоединение АТФ уменьшает сродство миозиновой головки с актиновыми нитями и миозин возвращается в исходное состояние. Далее повторяется весь цикл с самого начала, но поскольку в предыдущем цикле актиновая нить за счет своего движения приблизила Z-пластинку, то та же самая головка миозина присоединяется уже к другому мономеру актина ближе к Z-пластинке.
Сотни миозиновых головок каждой миозиновой нити работают одновременно, втягивая таким образом актиновую нить.
Источники энергии мышечного сокращения.
Скелетная мышца, работающая с максимальной интенсивностью, потребляет в сотни раз больше энергии, чем покоящаяся, причем переход от состояния покоя к состоянию максимальной работы происходит за доли секунды. В связи с этим у мышц совсем по-другому построен механизм изменения скорости синтеза АТФ в очень широких пределах.
Как уже упоминалось при мышечном сокращении большое значение имеет процесс синтеза АТФ из АДФ, высвобождаемых из миозиновых головок. Это происходит при помощи, имеющегося в мышцах высокоэнергетического вещества креатинфосфата , которое образуется из креатина и АТФ при действии креатинкиназы :
C-NH 2 C-NH-PO 3 H 2
N-CH 3 +АТФ- N-CH 3 + АДФ
Креатин Креатинфосфат
Эта реакция легко обратима и идет анаэробно, что обеспечивает быстрое включение мышц в работу на ранних этапах. При продолжении нагрузки роль такого энергетического обеспечения снижается, а на его замену приходят гликогеновые механизмы обеспечения большим количеством АТФ.
Библиография:
Г. Дюга, К. Пенни «Биоорганическая химия», М., 1983
Д. Мецлер «Биохимия», М., 1980
А. Ленинджер «Основы биохимии», М., 1985
БИОХИМИЯ МЫШЦ И МЫШЕЧНОГО СОКРАЩЕНИЯ. Механизм мышечного сокращения и расслабления. Важнейшей особенностью функционирования мышц является то что в процессе мышечного сокращения происходит непосредственное превращение химической энергии АТФ в механическую энергию сокращения мышц. Биохимически они различаются механизмами энергетического обеспечения мышечного сокращения.
Поделитесь работой в социальных сетях
Если эта работа Вам не подошла внизу страницы есть список похожих работ. Так же Вы можете воспользоваться кнопкой поиск
Лекция 7. Тема: БИОХИМИЯ МЫШЦ И МЫШЕЧНОГО СОКРАЩЕНИЯ
Вопросы:
2. Строение миофибрилл.
1. Общая характеристика мышц. Строение мышечных клеток.
Учение о мышцах это важнейший раздел биохимии, имеющий исключительное значение для спортивной биохимии.
Важнейшей особенностью функционирования мышц является то, что в процессе мышечного сокращения происходит непосредственное превращение химической энергии АТФ в механическую энергию сокращения мышц. Это явление не имеет аналогов в технике и присуще только живым организмам.
При изучении скелетных мышц с помощью светового микроскопа в них обнаружили поперечную исчерченность; отсюда их название поперечно-полосатые.
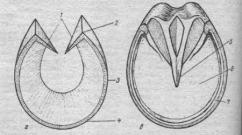
В скелетной мышце выделяют сухожильную головку, которой мышца начинается на кости, мышечное брюшко, состоящее из волокон, и сухожильный хвост, которым мышца заканчивается на другой кости (рис.).
Мышечное волокно структурная единица мышцы. Известны три типа мышечных волокон: белые быстро сокращающиеся ( VT ), промежуточные ( FR ) и медленно сокращающиеся ( ST ). Биохимически они различаются механизмами энергетического обеспечения мышечного сокращения. Их иннервируют разные мотонейроны, чем обусловлены неодновременность включения в работу и различная скорость сокращения волокон. Разные мышцы имеют разное сочетание типов волокон.
Мышечные волокна
Сухожилие
Рисунок. Мышца
Каждая мышца состоит из нескольких тысяч мышечных волокон, объединяемых соединительными прослойками и такой же оболочкой. Мышца представляет собой многокомпонентный комплекс. Чтобы разобраться в строении мышцы следует изучить все уровни ее организации и структуры, входящие в ее состав.
У животных и человека два основных типа мышц: поперечнополосатые и гладкие , причем поперечнополосатые мышцы делятся на два вида скелетные и сердечные . Гладкие мышцы характерны для внутренних органов, кровеносных сосудов.
Поперечнополосатые мышцы состоят из тысяч мышечных клеток волокон. Волокна объединены соединительно-тканными прослойками и такой же оболочкой фасцией . Мышечные волокна миоциты - представляют собой сильно вытянутые многоядерные клетки гигантских размеров от 0,1 до 10см длиной и толщиной около 0,1 0,2 мм.
Миоцит состоит из всех обязательных компонентов клетки. Особенностью мышечного волокна является то, что внутри эта клетка содержит большое количество сократительных элементов миофибрилл. Как и другие клетки тела миоциты содержат ядро, причем, у клеток поперечнополосатых мышц ядер несколько, рибосомы, митохондрии, лизосомы, цитоплазматическую сеть.
Цитоплазматическая сеть называется в этих клетках саркоплазматической сетью. Она связана с помощью особых трубочек, называемых Т-трубочками, с клеточной мембранной сарколеммой. Особо следует выделить в саркоплазматической сети пузырьки, называемые цистернами. Они содержат большое количество ионов кальция. С помощью специального фермента кальций накачивается в цистерны. Этот механизм называется кальциевым насосом и необходим для сокращения мышцы.
Цитоплазма или саркоплазма миоцитов содержит большое количество белков. Здесь немало активных ферментов, среди которых важнейшими являются ферменты гликолиза , креатинкиназа. Немалое значение имеет белок миоглобин, сохраняющий кислород в мышцах.
Кроме белков в цитоплазме мышечных клеток содержатся фосфогены АТФ, АДФ, АМФ, а также креатинфосфат, необходимые для нормального снабжения мышцы энергией.
Основной углевод мышечной ткани гликоген. Его концентрация достигает 3%. Свободная глюкоза в саркоплазме встречается в малых концентрациях. В тренируемых на выносливость мышцах накапливается запасной жир .
Снаружи сарколемма окружена нитями белка коллагена. Мышечное волокно растягивается и возвращается в исходное состояние за счет упругих сил, возникающих в коллагеновой оболочке.
2. Строение миофибрилл.
Сократительные элементы миофибриллы занимают большую часть объема миоцитов. В нетренированных мышцах миофибриллы расположены, рассеяно, а тренированных они сгруппированы в пучки, называемые полями Конгейма .
Микроскопическое изучение строения миофибрилл показало, что они имеют диаметр около 1 мкм и состоят из чередующихся светлых и темных участков или дисков. В мышечных клетках миофибриллы располагаются таким образом, что светлые и темные участки рядом расположенных миофибрилл совпадают, что создает видимую под микроскопом поперечную исчертанность всего мышечного волокна.
Использование электронного микроскопа с очень большим увеличением позволило расшифровать строение миофибрилл и установить причины наличия у них светлых и темных участков. Было обнаружено, что миофибриллы являются сложными структурами, построенными в свою очередь, из большого числа мышечных нитей дух типов толстых и тонких. Толстые в два раза толще тонких, соответственно 15 и 7 нм.
Состоят миофибриллы из чередующихся пучков параллельно расположенных толстых и тонких нитей, которые концами заходят друг на друга.
Участок миофибриллы, состоящий из толстых нитей и находящимися между ними концов тонких нитей, обладает двойным лучепреломлением. Под микроскопом эти участки кажутся темными и получили название анизотропных или темных дисков (А-диски).
Тонкие участки состоят из тонких нитей и выглядят светлыми, так как не обладают двойным лучепреломлением и легко пропускают свет. Такие участки называются изотропными или светлыми дисками ( I -диски).
Z Z Z
I -диск А-диск
Рисунок. Схема строения миофибриллы
В середине пучка тонких нитей (диск I ) поперечно располагается тонкая пластинка из белка, которая фиксирует положение мышечных нитей в пространстве и одновременно упорядочивая расположение А- и I -дисков многих миофибрилл. Эта пластинка хорошо видна под микроскопом и названа Z -пластинкой или Z -линией .
Диски А имеют в середине более светлую полосу зону Н, пересекаемую более темной М зоной.
Участок между соседними Z -линиями называется саркомер. Каждая миофибрилла состоит из нескольких сотен саркомеров (до 1000-1200).
саркомер
а

I -диск А-диск I -диск
Рисунок. Структура мышцы на разных уровнях организации: а мышечное волокно; б расположение миофибриллы в покоящейся мышце
Каждый саркомер включает: 1) сеть поперечных трубочек, ориентированных под углом 90° к продольной оси волокна и соединяющихся с наружной поверхностью клетки; 2) саркоплаз-матический ретикулум, составляющий 810% объема клетки; 3) несколько митохондрий.
Диски I состоят только из тонких филаментов, а диски А из филаментов двух типов. Зона Н содержит только толстые филаменты, линия Z скрепляет тонкие филаменты между собой. Между толстыми и тонкими филаментами расположены поперечные мостики (спайки) толщиной около 3 нм; расстояние между этими мостиками 40 нм.
Изучение химического состава миофибрилл показало, что тонкие и толстые нити образованы белками . Палочковидная молекула миозина состоит из двух идентичных основных цепей (по 200 кДа) и четырех легких цепей (по 20 кДа), общая масса миозина около 500 кДа.
Толстые нити состоят из белка миозина. Эти белки образуют двойную спираль с глобулярной головкой на конце, присоединенной к очень длинному стержню. Стержень представляет собой двухцепочечную а-спирализованную суперспираль.
Миозиновые головки обладают АТФазной активностью, то есть способностью расщеплять АТФ. Второй участок миозина обеспечивает связь толстых нитей с тонкими. Общая структура миозина показана на рисунке.
хвост

Рисунок. Схематичное изображение молекулы миозина
Тонкие нити состоят из белков актина, тропонина и тропомиозина.
Основной белок в данном случае актин . Он обладает двумя важнейшими свойствами:
- образует фибриллярный актин, способный к быстрой полимеризации;
- актин способен соединяться с миозиновыми головками поперечными мостиками.
Актин водорастворимый глобулярный белок с молекулярной массой 42 кДа; эта форма актина обозначается как G -актин. В мышечном волокне актин находится в полимеризованной форме, которая обозначается как F -актин. Тонкие филаменты мышцы образованы двунитчатыми актиновыми структурами, связанными между собой нековалентными связями.
Другие белки тонких нитей помогают актину осуществлять его функции.
Тропонин (Тн), молекулярная масса которого около 76 кДа. Он представляет собой сферическую молекулу, состоящую из трех разных субъединиц, получивших название в соответствии с выполняемыми функциями: тропомиозинсвязывающей (Тн-Т), ингибирующей (Тн-1) и кальцийсвязывающей (Тн-С). Каждый компонент тонких филаментов соединяется с двумя другими нековалентными связями:
F
-актин тропомиозин
Тн-1
Тн-Т
В мышце, где все рассмотренные компоненты собраны вместе в тонком филаменте (рис.), тропомиозин блокирует присоединение миозиновой головки к находящемуся рядом молекулами глобулярного актина тонких нитей ( F -актину).
Молекулы миозина объединяются, образуя филаменты, состоящие примерно из 400 палочковидных молекул, связанных друг с другом таким образом, что пары головок миозиновых молекул ложатся на расстоянии 14,3 нм друг от друга; они располагаются по спирали (рис.). Миозиновые нити стыкуются «хвост к хвосту».

Рисунок. Упаковка миозиновых молекул при образовании толстого филамента
Миозин выполняет три биологически важные функции:
При физиологических значениях ионной силы и рН молекулы миозина спонтанно образуют волокно.
Миозин обладает каталитической активностью, т. е. является ферментом. В 1939 г. ВА Энгельгардт и М.Н. Любимова обнаружили, что миозин способен катализировать гидролиз АТФ. Эта реакция является непосредственным источником свободной энергии, необходимой для мышечного сокращения.
Миозин связывает полимеризованную форму актина основного белкового компонента тонких миофибрилл. Именно это взаимодействие, как будет показано ниже, играет ключевую роль в мышечном сокращении.
Строение и механизм сокращения скелетных мышц.

3. Механизм мышечного сокращения и расслабления.
Подвижность является характерным свойством всех форм жизни. Направленное движение имеет место при расхождении хромосом в процессе клеточного деления, активном транспорте молекул, перемещении рибосом в ходе белкового синтеза, сокращении и расслаблении мышц. Мышечное сокращение наиболее совершенная форма биологической подвижности. В основе любого движения, в том числе и мышечного, лежат общие молекулярные механизмы.
У человека различают несколько видов мышечной ткани. Поперечно-полосатая мышечная ткань составляет мышцы скелета (скелетные мышцы, которые мы можем сокращать произвольно). Гладкая мышечная ткань входит в состав мышц внутренних органов: желудочно-кишечного тракта, бронхов, мочевыводящих путей, кровеносных сосудов. Эти мышцы сокращаются непроизвольно, независимо от нашего сознания.
В данной главе мы рассмотрим строение и процессы сокращения и расслабления скелетных мышц, поскольку именно они представляют наибольший интерес для биохимии спорта.
Механизм мышечного сокращения до настоящего времени раскрыт не полностью.
Достоверно известно следующее.
1. Источником энергии для мышечного сокращения являются молекулы АТФ.
2. Гидролиз АТФ катализируется при мышечном сокращении миозином, обладающим ферментативной активностью.
3. Пусковым механизмом мышечного сокращения является повышение концентрации ионов кальция в саркоплазме миоцитов, вызываемое нервным двигательным импульсом.
4. Во время мышечного сокращения между тонкими и толстыми нитями миофибрилл возникают поперечные мостики или спайки.
5. Во время мышечного сокращения происходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом.
Гипотез объясняющих механизм мышечного сокращения много, но наиболее обоснованной является так называемая гипотеза (теория) «скользящих нитей» или «гребная гипотеза».
В покоящейся мышце тонкие и толстые нити находятся в разъединенном состоянии.
Под воздействием нервного импульса ионы кальция выходят из цистерн саркоплазматической сети и присоединяются к белку тонких нитей тропонину. Этот белок меняет свою конфигурацию и меняет конфигурацию актина. В результате образуется поперечный мостик между актином тонких нитей и миозином толстых нитей. При этом повышается АТФазная активность миозина. Миозин расщепляет АТФ и за счет выделившейся при этом энергии миозиновая головка подобно шарниру или веслу лодки поворачивается, что приводит к скольжению мышечных нитей навстречу друг другу.
Совершив поворот, мостики между нитями разрываются. АТФазная активность миозина резко снижается, прекращается гидролиз АТФ. Однако при дальнейшем поступлении нервного импульса поперечные мостики вновь образуются, так как процесс, описанный выше, повторяется вновь.
В каждом цикле сокращения расходуется 1 молекула АТФ.
В основе мышечного сокращения лежат два процесса:
спиральное скручивание сократительных белков;
циклически повторяющееся образование и диссоциация комплекса между цепью миозина и актином.
Мышечное сокращение инициируется приходом потенциала действия на концевую пластинку двигательного нерва, где выделяется нейрогормон ацетилхолин, функцией которого является передача импульсов. Сначала ацетилхолин взаимодействует с ацетилхолиновыми рецепторами, что приводит к распространению потенциала действия вдоль сарколеммы. Все это вызывает увеличение проницаемости сарколеммы для катионов Na + , которые устремляются внутрь мышечного волокна, нейтрализуя отрицательный заряд на внутренней поверхности сарколеммы. С сарколеммой связаны поперечные трубочки саркоплазматического ретикулума, по которым распространяется волна возбуждения. От трубочек волна возбуждения передается мембранам пузырьков и цистерн, которые оплетают миофибриллы на участках, где происходит взаимодействие актиновых и миозиновых нитей. При передаче сигнала на цистерны саркоплазматического ретикулума, последние начинают освобождать находящийся в них Са 2+ . Высвобожденный Са 2+ связывается с Тн-С, что вызывает конформационные сдвиги, передающиеся на тропомиозин и далее на актин. Актин как бы освобождается из комплекса с компонентами тонких филамен-тов, в котором он находился. Далее актин взаимодействует с миозином, и результатом такого взаимодействия является образование спайки, что делает возможным движение тонких нитей вдоль толстых.
Генерация силы (укорочение) обусловлена характером взаимодействия между миозином и актином. На миозиновом стержне имеется подвижный шарнир, в области которого происходит поворот при связывании глобулярной головки миозина с определенным участком актина. Именно такие повороты, происходящие одновременно в многочисленных участках взаимодействия миозина и актина, являются причиной втягивания актиновых филаментов (тонких нитей) в Н-зону. Здесь они контактируют (при максимальном укорочении) или даже перекрываются друг с другом, как это показано на рисунке.


б
 в
в
Рисунок. Механизм сокращения: а состояние покоя; б умеренное сокращение; в максимальное сокращение
Энергию для этого процесса поставляет гидролиз АТФ. Когда АТФ присоединяется к головке молекулы миозина, где локализован активный центр миозиновой АТФазы, связи между тонкой и толстой нитями не образуется. Появившийся катион кальция нейтрализует отрицательный заряд АТФ, способствуя сближению с активным центром миозиновой АТФазы. В результате происходит фосфорилирование миозина, т. е. миозин заряжается энергией, которая используется для образования спайки с актином и для продвижения тонкой нити. После того как тонкая нить продвинется на один «шаг», АДФ и фосфорная кислота отщепляются от актомиозинового комплекса. Затем к миозиновой головке присоединяется новая молекула АТФ, и весь процесс повторяется со следующей головкой молекулы миозина.
Затрата АТФ необходима и для расслабления мышц. После прекращения действия двигательного импульса Са 2+ переходит в цистерны саркоплазматического ретикулума. Тн-С теряет связанный с ним кальций, следствием этого являются конформаци-онные сдвиги в комплексе тропонин-тропомиозин, и Тн- I снова закрывает активные центры актина, делая их неспособными взаимодействовать с миозином. Концентрация Са 2+ в области сократительных белков становится ниже пороговой, и мышечные волокна теряют способность образовывать актомиозин.
В этих условиях эластические силы стромы, деформированной в момент сокращения, берут верх, и мышца расслабляется. При этом тонкие нити извлекаются из пространства между толстыми нитями диска А, зона Н и диск I приобретают первоначальную длину, линии Z отдаляются друг от друга на прежнее расстояние. Мышца становится тоньше и длиннее.
Скорость гидролиза АТФ при мышечной работе огромна: до 10 мк моль на 1 г мышцы за 1 мин. Общие запасы АТФ невелики, поэтому для обеспечения нормальной работы мышц АТФ должна восстанавливаться с той же скоростью, с какой она расходуется.
Расслабление мышцы происходит после прекращения поступления длительного нервного импульса. При этом проницаемость стенки цистерн саркоплазматической сети уменьшается, и ионы кальция под действием кальциевого насоса, используя энергию АТФ, уходят в цистерны. Концентрация ионов кальция в саркоплазме быстро снижается до исходного уровня.Белки вновь приобретают конформацию характерную для состояния покоя.
Таким образом, и процесс мышечного сокращения и процесс мышечного расслабления это активные процессы, идущие с затратами энергии в виде молекул АТФ,
В гладких мышцах нет миофибрилл. Тонкие нити присоединяются к сарколемме, толстые находятся внутри волокон. Ионы кальция также играют роль в сокращении, но поступают в мышцу не из цистерн, а из внеклеточного вещества, поскольку в гладких мышцах отсутствуют цистерны с ионами калькия. Этот процесс медленный и поэтому медленно работают гладкие мышцы.
Рисунок. Схема расположения толстых и тонких нией в гладких мышечных волокнах.
Другие похожие работы, которые могут вас заинтересовать.вшм> |
|||
| 379. | ЭНЕРГЕТИЧЕСКОЕ ОБЕСПЕЧЕНИЕ МЫШЕЧНОГО СОКРАЩЕНИЯ | 33.58 KB | |
| Количественные критерии путей ресинтеза АТФ. Аэробный путь ресинтеза АТФ. Анаэробные пути ресинтеза АТФ. Соотношения между различными путями ресинтеза АТФ при мышечной работе. | |||
| 17220. | Биохимия | 122.66 KB | |
| Опишите изменение обмена веществ в клетках-мишенях под действием инсулина укажите: характер влияния гормона на уровень циклических нуклеотидов; изменение проницаемости мембран для различных веществ; регулируемые биохимические процессы; конечный биологический эффект... | |||
| 21483. | БИОХИМИЯ ГОРМОНОВ | 63.62 KB | |
| Гормоны являются первичными посредниками между центральной нервной системой и тканевыми процессами. Термин гормоны 1905 году ввели ученые Бейлис и Старлинг. Их называют по месту образования инсулин от insul-островок по физиологическому эффекту вазопрессин гормоны передней доли гипофиза имеют окончание тропин окончание либерин и статин указывает на гипоталамические гормоны. | |||
| 21608. | БИОХИМИЯ КРОВИ | 95.89 KB | |
| Гемоглобин по своей химической природе относится к гемопротеидам, состоит из простатической группы гема и белка глобина. Гем - тетрапирольное железосодержащее органическое вещество. Гем соединяется с гемоглобином гидрофобными связями и координационной связью с железом. Гемоглобин является олигомерным белком, включает в свой состав 4 гема и 4 полипептидные цепи. | |||
| 10034. | Пути сокращения производственных запасов | 106.84 KB | |
| На сегодняшний момент времени, главная задача предприятий - значительное повышение качества производственного процесса, его эффективности, отдачи вложений, в том числе и производственных, которые являются базой всего производства. | |||
| 15050. | Пути сокращения затрат предприятия ООО «Томак-2» | 138.77 KB | |
| Проблемы снижения затрат на предприятии, поиска путей их решения являются сложными и интересными вопросами современной экономики предприятия. Проблема снижения затрат очень актуальна в современных экономических условиях, так как ее решение позволяет каждому конкретному предприятию выжить в условиях жесткой рыночной конкуренции, построить крепкое и сильное предприятие, которое будет иметь хороший экономический потенциал. | |||
| 5067. | Гладкие мышцы. Строение, функции, механизм сокращения | 134.79 KB | |
| Мышцы или мускулы от лат. Мышцы позволяют двигать частями тела и выражать в действиях мысли и чувства. Гладкие мышцы являются составной частью некоторых внутренних органов и участвуют в обеспечении функции выполняемые этими органами. | |||
| 17984. | Перспективы сокращения и социально-экономическое значение государственного долга Российской Федерации | 395.55 KB | |
| Причины возникновения государственного долга Российской Федерации. Анализ и современное состояние государственного внутреннего долга Российской Федерации. Анализ и современное состояние государственного внешнего долга Российской Федерации. Перспективы сокращения и социально-экономическое значение государственного долга Российской Федерации... | |||
| 11490. | Пути сокращения длительности товарооборота предприятий розничной торговли (на материалах ООО «Диана», г. Курган) | 176.54 KB | |
| Размер товарных запасов является синтетическим показателем, позволяющим в известной мере оценивать результаты хозяйственной деятельности, как отдельных торговых предприятий, организаций, так и отрасли в целом, а также эффективность использования материальных и трудовых ресурсов. | |||
| 12159. | О стратегической стабильности в прошлом и настоящем и ее значении для выработки подходов к ограничению и сокращения вооружений | 17.33 KB | |
| Проведен анализ угроз стратегической стабильности сформировавшихся за последние годы прежде всего за счет распространения ядерного оружия. Показано что стратегическая стабильность в большей степени чем прежде зависит от нарушения региональной стабильности. Проблема обеспечения ядерной стабильности остается актуальной и для диадных отношений РоссияСША. | |||
Легкий мирозин отличается от тяжелого по аминокислотному составу свойствами. Тяжелый миозин обладает ферментативной активностью. Он является аденозинтрифосфотазой и гидролитически расщепляет АТФ. Это можно описать: АТФ + H 2 O → АДФ + H 3 PO 4 + W (энергия).
Актин – белок с более низкой молекулярной массой (42000). Может быть в двух формах: глобулярной ( G ) или фибриллярной ( F ). После прибавления солей G -актин легко переходит в F -актин. F -актин является полимером G -актина. Этот переход осуществляется под влиянием ионов К + : актин глобулярный → акт ин фибрилярный F . Актин F легко соединяется с миозином и образует новый белок-актомиозин.
F -актин состоит из двух филаментов скрученных в спираль.
Структура актина
Для актомиозина характерны следующие свойства:
способность разлагать АТФ;
освобождать энергию макроэргических связей;
превращать эту энергию в работу.
Тропомиозин – состоит из двух полипептидных цепей образующих двойную спираль, располагается в бороздке на поверхности – F актина по длине соответствует 7 субъектам - G -актина. Комплекс тропонина состоит из трех субъединиц с глобулярной структурой и расположен примерно на концах Т m . Tропонин Т ( TnT ) обеспечивает связь с Т m . T ропонин С ( TnC ) образует связь с ионами Са 2+ на поверхности Т m , в результате чего изменяется его конформация.
Тропонин I ( TnI ) может предотвращать взаимодействие актина с миозином. Положение Т nI переменно и зависит от концентрации Са 2+ . В присутствии Са 2+ изменяется конформация Т nC . Это приводит к изменению положения TnI по отношению к актину, в результате он может взаимодействовать с миозином.
Тропомиозин и тропонин
Точное
пространственное расположение главных
белков сократительной мышцы - необходимое
условие сокращения и расслабления, а
также регуляции этих процессов. Сокращение
связано с образованием комплекса между
актином и миозином, в котором каждая
субъединица актина взаимодействует с
сегментом, содержащим головку миозина
(
F
1
).
Расслабление происходит при сокращении
этого взаимодействия. Взаимодействие
А и М регулируется T, который находится
в бороздке актина. Изменение конформации
Т передается на T, который погружается
глубже в бороздку разрешая взаимодействие
актина с головкой миозина.

Состояние миофибриллы: а) покой; б) сокращение
Миоглобин – сложный белок хромопротеин, по строению близок к гемоглобину, находится в красных мышцах, способен связывать и отдавать кислород, способствуя снабжению мышечных волокон кислородом.
В состав белков протоплазмы входят ферменты гликолиза с высокой ферментативной активностью. Ферменты биологического окисления сосредоточены в митохондриях, где осуществляется окислительное фосфорилирование. В рибосомах, лизосомах содержатся ферменты, осуществляющие превращение белков и липидов.
Оксимиоглобин отдает кислород только при значительном снижении парциального давления. Миоглобин извлекается из тканей аммиачным раствором. Соединительнотканные белки входят в состав оболочек клетки и субклеточных образований, стенок сосудов, нервов. Их содержание составляет до 20% от общего количества мышц. Это главным образом коллаген; они не извлекаются даже растворами солей.
В мышце имеются аминокислоты, полипептиды, а также азотосодержащие вещества, которые легко извлекаются водой. Их называют экстрактивными веществами. К ним относятся креатин и креатинфосфат, на долю которых приходится до 60% всего небелкового азота. В покое весь креатин мышц представлен в виде креатинфосфата. Его концентрация в мышце довольно высокая (0,2-0,55%), в связи с тем, что он играет важную роль в передаче макроэргических связей внутри клетки, и обеспечивает ресинтез АТФ.
Креатинфосфат (КрФ) - макроэргическое соединение, способное отдавать фосфорную группу на АДФ; реакцию катализирует креатинфосфаткиназа по схеме:
АДФ + Крф → креатинфосфаткиназа АТФ – Кр ( креатин )

Креатин синтезируется в почках из аргинина.

В мышцы креатин доставляется с кровью.
Креатинфосфат (Крф) резерв макроэргических связей в мышце.

В мышцах можно обнаружить и некоторое количество креатинина, образующегося при разрушении Крф (креатинфосфата).
К числу азотосодержащих экстрактивных веществ относится ансерин, карнитин, карнозин (β-аланин-гистидин). В мышцах высоко содержание адениловых нуклеотидов, которые относятся к экстрактивным веществам (до 0,4 %) АТФ, АМФ, АДФ.
Углеводы представлены в основном гликогеном (0,5-0,8%). Основная масса гликогена организма сосредоточена в мышцах, хотя в печени его концентрация выше (5%). Моносахариды представлены преимущественно в виде гексозофосфатов, их концентрация не превышает концентрацию глюкозы в крови.
Минеральные вещества – (зола) составляет 1-1,5% массы мышц. Наряду с К + и Na + в мышцах содержится Ca 2+ и М g 2+ , которые играют важную роль в механизме мышечного сокращения. В условиях покоя Са 2+ сосредоточен преимущественно в трубочках и пузырьках саркоплазматического ретикулума.
Основная масса фосфора (около 80%) мышечной ткани входит в состав макроэргических соединений (АТФ и креатинфосфат), 10% представлено в виде солей неорганического фосфата, 5% связано с гексозами и 5% входит в состав АДФ, АМФ и других нуклеотидов.
Химический состав гладких мышц включает те же вещества, что и поперечно-полосатые мышцы, но в других количественных соотношениях. В них меньше актомиозина и миозина, но больше миоальбумина и нерастворимых белков стромы (коллагена). Содержание гликогена менее 0,5%, меньше и экстрактивных веществ. Содержание Са 2+ в гладких мышцах ниже.
Подвижность является характерным свойством всех форм жизни. Направленное движение имеет место при расхождении хромосом в процессе клеточного деления, активном транспорте молекул, перемещении рибосом в ходе белкового синтеза, сокращении и расслаблении мышц. Мышечное сокращение – наиболее совершенная форма биологической подвижности. В основе любого движения, в том числе и мышечного, лежат общие молекулярные механизмы.
У человека различают несколько видов мышечной ткани. Поперечно-полосатая мышечная ткань составляет мышцы скелета (скелетные мышцы, которые мы можем сокращать произвольно). Гладкая мышечная ткань входит в состав мышц внутренних органов: желудочно-кишечного тракта, бронхов, мочевыводящих путей, кровеносных сосудов. Эти мышцы сокращаются непроизвольно, независимо от нашего сознания.
В данной лекции мы рассмотрим строение и процессы сокращения и расслабления скелетных мышц, поскольку именно они представляют наибольший интерес для биохимии спорта.
Механизм мышечного сокращения до настоящего времени раскрыт не полностью.
Достоверно известно следующее.
1. Источником энергии для мышечного сокращения являются молекулы АТФ.
2. Гидролиз АТФ катализируется при мышечном сокращении миозином, обладающим ферментативной активностью.
3. Пусковым механизмом мышечного сокращения является повышение концентрации ионов кальция в саркоплазме миоцитов, вызываемое нервным двигательным импульсом.
4. Во время мышечного сокращения между тонкими и толстыми нитями миофибрилл возникают поперечные мостики или спайки.
5. Во время мышечного сокращения происходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом.
Гипотез объясняющих механизм мышечного сокращения много, но наиболее обоснованной является так называемая гипотеза (теория) «скользящих нитей» или «гребная гипотеза».
В покоящейся мышце тонкие и толстые нити находятся в разъединенном состоянии.
Под воздействием нервного импульса ионы кальция выходят из цистерн саркоплазматической сети и присоединяются к белку тонких нитей – тропонину. Этот белок меняет свою конфигурацию и меняет конфигурацию актина. В результате образуется поперечный мостик между актином тонких нитей и миозином толстых нитей. При этом повышается АТФазная активность миозина. Миозин расщепляет АТФ и за счет выделившейся при этом энергии миозиновая головка подобно шарниру или веслу лодки поворачивается, что приводит к скольжению мышечных нитей навстречу друг другу.
Совершив поворот, мостики между нитями разрываются. АТФазная активность миозина резко снижается, прекращается гидролиз АТФ. Однако при дальнейшем поступлении нервного импульса поперечные мостики вновь образуются, так как процесс, описанный выше, повторяется вновь.
В каждом цикле сокращения расходуется 1 молекула АТФ.
В основе мышечного сокращения лежат два процесса:
Спиральное скручивание сократительных белков;
Циклически повторяющееся образование и диссоциация комплекса между цепью миозина и актином.
Мышечное сокращение инициируется приходом потенциала действия на концевую пластинку двигательного нерва, где выделяется нейрогормон ацетилхолин, функцией которого является передача импульсов. Сначала ацетилхолин взаимодействует с ацетилхолиновыми рецепторами, что приводит к распространению потенциала действия вдоль сарколеммы. Все это вызывает увеличение проницаемости сарколеммы для катионов Na + , которые устремляются внутрь мышечного волокна, нейтрализуя отрицательный заряд на внутренней поверхности сарколеммы. С сарколеммой связаны поперечные трубочки саркоплазматического ретикулума, по которым распространяется волна возбуждения. От трубочек волна возбуждения передается мембранам пузырьков и цистерн, которые оплетают миофибриллы на участках, где происходит взаимодействие актиновых и миозиновых нитей. При передаче сигнала на цистерны саркоплазматического ретикулума, последние начинают освобождать находящийся в них Са 2+ . Высвобожденный Са 2+ связывается с Тн-С, что вызывает конформационные сдвиги, передающиеся на тропомиозин и далее на актин. Актин как бы освобождается из комплекса с компонентами тонких филаментов, в котором он находился. Далее актин взаимодействует с миозином, и результатом такого взаимодействия является образование спайки, что делает возможным движение тонких нитей вдоль толстых.
Генерация силы (укорочение) обусловлена характером взаимодействия между миозином и актином. На миозиновом стержне имеется подвижный шарнир, в области которого происходит поворот при связывании глобулярной головки миозина с определенным участком актина. Именно такие повороты, происходящие одновременно в многочисленных участках взаимодействия миозина и актина, являются причиной втягивания актиновых филаментов (тонких нитей) в Н-зону. Здесь они контактируют (при максимальном укорочении) или даже перекрываются друг с другом, как это показано на рисунке.

 в
в
Рисунок. Механизм сокращения: а – состояние покоя; б – умеренное сокращение; в – максимальное сокращение
Энергию для этого процесса поставляет гидролиз АТФ. Когда АТФ присоединяется к головке молекулы миозина, где локализован активный центр миозиновой АТФазы, связи между тонкой и толстой нитями не образуется. Появившийся катион кальция нейтрализует отрицательный заряд АТФ, способствуя сближению с активным центром миозиновой АТФазы. В результате происходит фосфорилирование миозина, т. е. миозин заряжается энергией, которая используется для образования спайки с актином и для продвижения тонкой нити. После того как тонкая нить продвинется на один «шаг», АДФ и фосфорная кислота отщепляются от актомиозинового комплекса. Затем к миозиновой головке присоединяется новая молекула АТФ, и весь процесс повторяется со следующей головкой молекулы миозина.
Затрата АТФ необходима и для расслабления мышц. После прекращения действия двигательного импульса Са 2+ переходит в цистерны саркоплазматического ретикулума. Тн-С теряет связанный с ним кальций, следствием этого являются конформаци-онные сдвиги в комплексе тропонин-тропомиозин, и Тн-I снова закрывает активные центры актина, делая их неспособными взаимодействовать с миозином. Концентрация Са 2+ в области сократительных белков становится ниже пороговой, и мышечные волокна теряют способность образовывать актомиозин.
В этих условиях эластические силы стромы, деформированной в момент сокращения, берут верх, и мышца расслабляется. При этом тонкие нити извлекаются из пространства между толстыми нитями диска А, зона Н и диск I приобретают первоначальную длину, линии Z отдаляются друг от друга на прежнее расстояние. Мышца становится тоньше и длиннее.
Скорость гидролиза АТФ при мышечной работе огромна: до 10 мк моль на 1 г мышцы за 1 мин. Общие запасы АТФ невелики, поэтому для обеспечения нормальной работы мышц АТФ должна восстанавливаться с той же скоростью, с какой она расходуется.
Расслабление мышцы происходит после прекращения поступления длительного нервного импульса. При этом проницаемость стенки цистерн саркоплазматической сети уменьшается, и ионы кальция под действием кальциевого насоса, используя энергию АТФ, уходят в цистерны. Удаление ионов кальция в цистерны ретикулума после прекращения двигательного импульса требует значительных энерготрат. Так как удаление ионов кальция происходит в сторону более высокой концетрации, т.е. против осмотического градиента, то на удаление каждого иона кальция затрачивается две молекулы АТФ. Концентрация ионов кальция в саркоплазме быстро снижается до исходного уровня. Белки вновь приобретают конформацию характерную для состояния покоя.
Таким образом, и процесс мышечного сокращения и процесс мышечного расслабления – это активные процессы, идущие с затратами энергии в виде молекул АТФ,
В гладких мышцах нет миофибрилл, которые состоят из нескольких сотен саркомеров. Тонкие нити присоединяются к сарколемме, толстые находятся внутри волокон. Ионы кальция также играют роль в сокращении, но поступают в мышцу не из цистерн, а из внеклеточного вещества, поскольку в гладких мышцах отсутствуют цистерны с ионами калькия. Этот процесс медленный и поэтому медленно работают гладкие мышцы.
Тема 9. Биохимия мышечного сокращения.Тема 10. Энергетическое обеспечение мышечного сокращения.
Тема 9. БИОХИМИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ.
1. Общая характеристика мышц. Строение мышечных клеток.
2. Строение миофибрилл.
3. Сокращение и расслабление мышцы.
§ 1. Общая характеристика мышц. Строение мышечных клеток.
Учение о мышцах – это важнейший раздел биохимии, имеющий исключительное значение для спортивной биохимии.
Важнейшей особенностью функционирования мышц является то, что в процессе мышечного сокращения происходит непосредственное превращение химической энергии АТФ в механическую энергию сокращения мышц. Это явление не имеет аналогов в технике и присуще только живым организмам.
У животных и человека два основных типа мышц: поперечнополосатые и гладкие , причем поперечнополосатые мышцы делятся на два вида – скелетные и сердечные . Гладкие мышцы характерны для внутренних органов, кровеносных сосудов.
Поперечнополосатые мышцы состоят из тысяч мышечных клеток – волокон. Волокна объединены соединительно-тканными прослойками и такой же оболочкой – фасцией. Мышечные волокна – миоциты - представляют собой сильно вытянутые многоядерные клетки гигантских размеров от 0,1 до 10см длиной и толщиной около 0,1 – 0,2 мм.
Миоцит состоит из всех обязательных компонентов клетки. Особенностью мышечного волокна является то, что внутри эта клетка содержит большое количество сократительных элементов - миофибрилл. Как и другие клетки тела миоциты содержат ядро, причем, у клеток поперечнополосатых мышц ядер несколько, рибосомы, митохондрии, лизосомы, цитоплазматическую сеть.
Цитоплазматическая сеть называется в этих клетках саркоплазматической сетью. Она связана с помощью особых трубочек, называемых Т-трубочками, с клеточной мембранной – сарколеммой. Особо следует выделить в саркоплазматической сети пузырьки, называемые цистернами. Они содержат большое количество ионов кальция. С помощью специального фермента кальций накачивается в цистерны. Этот механизм называется кальциевым насосом и необходим для сокращения мышцы.
Цитоплазма или саркоплазма миоцитов содержит большое количество белков. Здесь немало активных ферментов, среди которых важнейшими являются ферменты гликолиза , креатинкиназа. Немалое значение имеет белок миоглобин, сохраняющий кислород в мышцах.
Кроме белков в цитоплазме мышечных клеток содержатся фосфогены – АТФ, АДФ, АМФ, а также креатинфосфат, необходимые для нормального снабжения мышцы энергией.
Основной углевод мышечной ткани – гликоген. Его концентрация достигает 3%. Свободная глюкоза в саркоплазме встречается в малых концентрациях. В тренируемых на выносливость мышцах накапливается запасной жир .
Снаружи сарколемма окружена нитями белка – коллагена. Мышечное волокно растягивается и возвращается в исходное состояние за счет упругих сил, возникающих в коллагеновой оболочке.
§ 2. Сократительные элементы (миофибриллы ).
Сократительные элементы – миофибриллы – занимают большую часть объема миоцитов. В нетренированных мышцах миофибриллы расположены, рассеяно, а тренированных они сгруппированы в пучки, называемые полями Конгейма .
Микроскопическое изучение строения миофибрилл показало, что они состоят из чередующихся светлых и темных участков или дисков. В мышечных клетках миофибриллы располагаются таким образом, что светлые и темные участки рядом расположенных миофибрилл совпадают, что создает видимую под микроскопом поперечную исчертанность всего мышечного волокна.
Использование электронного микроскопа с очень большим увеличением позволило расшифровать строение миофибрилл и установить причины наличия у них светлых и темных участков. Было обнаружено, что миофибриллы являются сложными структурами, построенными в свою очередь, из большого числа мышечных нитей дух типов – толстых и тонких. Толстые в два раза толще тонких, соответственно 15 и 7 нм.
Состоят миофибриллы из чередующихся пучков параллельно расположенных толстых и тонких нитей, которые концами заходят друг на друга.
Участок миофибриллы, состоящий из толстых нитей и находящимися между ними концов тонких нитей, обладает двойным лучепреломлением. Под микроскопом эти участки кажутся темными и получили название анизотропных или темных дисков (А-диски).
Тонкие участки состоят из тонких нитей и выглядят светлыми. Такие участки называются изотропными или светлыми дисками (I -диски). В середине пучка тонких нитей поперечно располагается тонкая пластинка из белка, которая фиксирует положение мышечных нитей в пространстве. Эта пластинка хорошо видна под микроскопом и названа Z -пластинкой или Z -линией .
Участок между соседними Z-линиями называется саркомер. Каждая миофибрилла состоит из тысяч саркомеров.
Изучение химического состава миофибрилл показало, что тонкие и толстые нити образованы белками.
Толстые нити состоят из белка миозина. Эти белки образуют двойную спираль с глобулярной головкой на конце. Миозиновые головки обладают АТФазной активностью, то есть способностью расщеплять АТФ . Второй участок миозина обеспечивает связь толстых нитей с тонкими.
Тонкие нити состоят из белков актина, тропонина и тропомиозина.
Основной белок в данном случае актин. Он обладает двумя важнейшими свойствами:
образует фибриллярный актин, способный к быстрой полимеризации;
актин способен соединяться с миозиновыми головками поперечными мостиками.
На странице 44 на рисунке А. подробно показано строение мышц.
Строение и механизм сокращения скелетных мышц.
§ 3. Механизм мышечного сокращения и расслабления.
Механизм мышечного сокращения
до настоящего времени раскрыт не полностью.
Достоверно известно следующее.
1. Источником энергии для мышечного сокращения являются молекулы АТФ.
2. Гидролиз АТФ катализируется при мышечном сокращении миозином, обладающим ферментативной активностью.
3. Пусковым механизмом мышечного сокращения является повышение концентрации ионов кальция в саркоплазме миоцитов, вызываемое нервным двигательным импульсом.
4. Во время мышечного сокращения между тонкими и толстыми нитями миофибрилл возникают поперечные мостики или спайки.
5. Во время мышечного сокращения происходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом.
Гипотез объясняющих механизм мышечного сокращения много, но наиболее обоснованной является так называемая гипотеза (теория) «скользящих нитей» или «гребная гипотеза».
В покоящейся мышце тонкие и толстые нити находятся в разъединенном состоянии.
Под воздействием нервного импульса ионы кальция выходят из цистерн саркоплазматической сети и присоединяются к белку тонких нитей – тропонину. Этот белок меняет свою конфигурацию и меняет конфигурацию актина. В результате образуется поперечный мостик между актином тонких нитей и миозином толстых нитей. При этом повышается АТФазная активность миозина. Миозин расщепляет АТФ и за счет выделившейся при этом энергии миозиновая головка подобно шарниру или веслу лодки поворачивается, что приводит к скольжению мышечных нитей навстречу друг другу.
Совершив поворот, мостики между нитями разрываются. АТФазная активность миозина резко снижается, прекращается гидролиз АТФ. Однако при дальнейшем поступлении нервного импульса поперечные мостики вновь образуются, так как процесс, описанный выше, повторяется вновь.